本博客由科研AI Agent实验室BenszResearch强力驱动!如何更快地访问本站?有需要可加电报群获得更多帮助。本博客用什么VPS?创作不易,请支持苯苯!推荐购买本博客的VIP喔,10元/年即可畅享所有VIP专属内容!
概述
Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife. full pdf remote; full html; full pdf local;
Huan Yan 1,2 † , Guocai Zhong 2 † , Guangwei Xu 2 , Wenhui He 2,3 , Zhiyi Jing 2 , Zhenchao Gao 1,2 , Yi Huang 2,3 , Yonghe Qi 2 , Bo Peng 2 , Haimin Wang 2 , Liran Fu 2,3 , Mei Song 2,3 , Pan Chen 2,3 , Wenqing Gao 2 , Bijie Ren 2 , Yinyan Sun 2 , Tao Cai 2 , Xiaofeng Feng 2 , Jianhua Sui 2 , Wenhui Li 2 *
1 Graduate program in School of Life Sciences, Peking University, Beijing, China;
2 National Institute of Biological Sciences, Beijing, China;
3 Graduate program in Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, China李文辉教授获得2022年未来科学大奖的原创性研究,他是这篇文章的第一通讯作者
- 人类乙型肝炎病毒 (HBV) 感染和HBV相关疾病仍然是主要的公共卫生问题。合并感染其卫星丁型肝炎病毒 (HDV) 的个体患有更严重的疾病。两种病毒的细胞进入是由 HBV包膜蛋白介导的。大包膜蛋白的前 S1 结构域是受体结合的关键决定因素。然而,受体的身份是未知的。
- 在这里,通过使用近零距离光交联和串联亲和纯化,我们揭示了pre-S1的受体结合区域与主要在肝脏中表达的多跨膜转运蛋白牛磺胆酸钠协同转运多肽 (NTCP) 特异性相互作用。
- 沉默 NTCP 抑制了 HBV 和 HDV 感染,而外源性 NTCP 表达使不敏感的肝癌细胞对这些病毒感染敏感。
- 此外,用人类对应物替换非功能性猴子 NTCP 的氨基酸 157-165 赋予其支持两种病毒感染的能力。
- 我们的研究结果表明,NTCP 是 HBV 和 HDV 的功能性受体。
概览
- 本文围绕 概述 展开详细讨论
- 包含 12 个主要章节内容
- 文末提供总结与展望
前言
其实《经典精读》第1期已经准备了很久,只是之前忙于其它事情,一直没完成。这次趁国庆,就暂时完结它吧!虽然自己读得也不深入,但作为一个非正式的个人学习文章,在对主体框架基本理清的前提下发表在博客上没什么不妥。以后读到什么新的内容,再补充也不迟(主要是最近都用前沿快讯系列划水,有点不好意思,嘿嘿)。
这是一篇精心挑选的经典研究,它有以下特点:
- 近期李教授因此文获得了2022年度的未来科学大奖,关注度很高
- 10年前的作品,离我们并不是很遥远,和现在的研究范式比较接近,参考价值较强
- 中国科学家的重大原创性研究,对于中国年轻学者有很强的激励和示范作用
- 博文成书时,正值新冠病毒在全世界范围肆虐;这篇研究涉及病毒和人类,具有大众认知的舆论基础
- 论证过程充分运用了分子生物学的经典研究范式;研究设计包含了优质研究的基本特点,值得学习
- 久经考验,历史上似乎只有一次方法学(qPCR引物序列)的勘误
整体上,我觉得此文比较一般的CNS文章的阅读难度要低一些,解析过程也相对轻松,有疑惑的点看看正文基本上可以猜个八九成。不过,我自己并不是病毒受体研究领域的学者专家,水平有限,有不当错误之处,还请各位大佬批评指正了!
背景
- HBV是一个影响范围极广的传染性疾病。HBV相关肝病是一个严重的公共卫生问题。
- HBV进入(HBV entry)是一个很重要的生物学问题,它的悬而未决在很大程度影响了HBV的研究进展,比如药物研发。
- HDV是HBV的卫星病毒,仅与HBV共存时可以繁殖。由于HDV含有完整的3个HBV包膜蛋白,HDV被认为是研究HBV entry问题上的良好样本。
- L蛋白和S蛋白对HBV相关肝病的发展是十分重要的,它们与病毒进入肝细胞的生物学过程扮演重要角色。
- 在HBV的包膜蛋白中,pre-S1结构域已经被发现是病毒entry过程的关键功能片段之一;但是肝细胞的受体则一直未能明确。
- 除类人猿(chimpanzees)外,目前仅发现一种叫树鼩(shù qú; treeshrew/tupaia, 图片来自wikipedia)的小型哺乳动物对HBV感染敏感,即可作为HBV/HDV感染性肝炎研究的模式生物。

结果
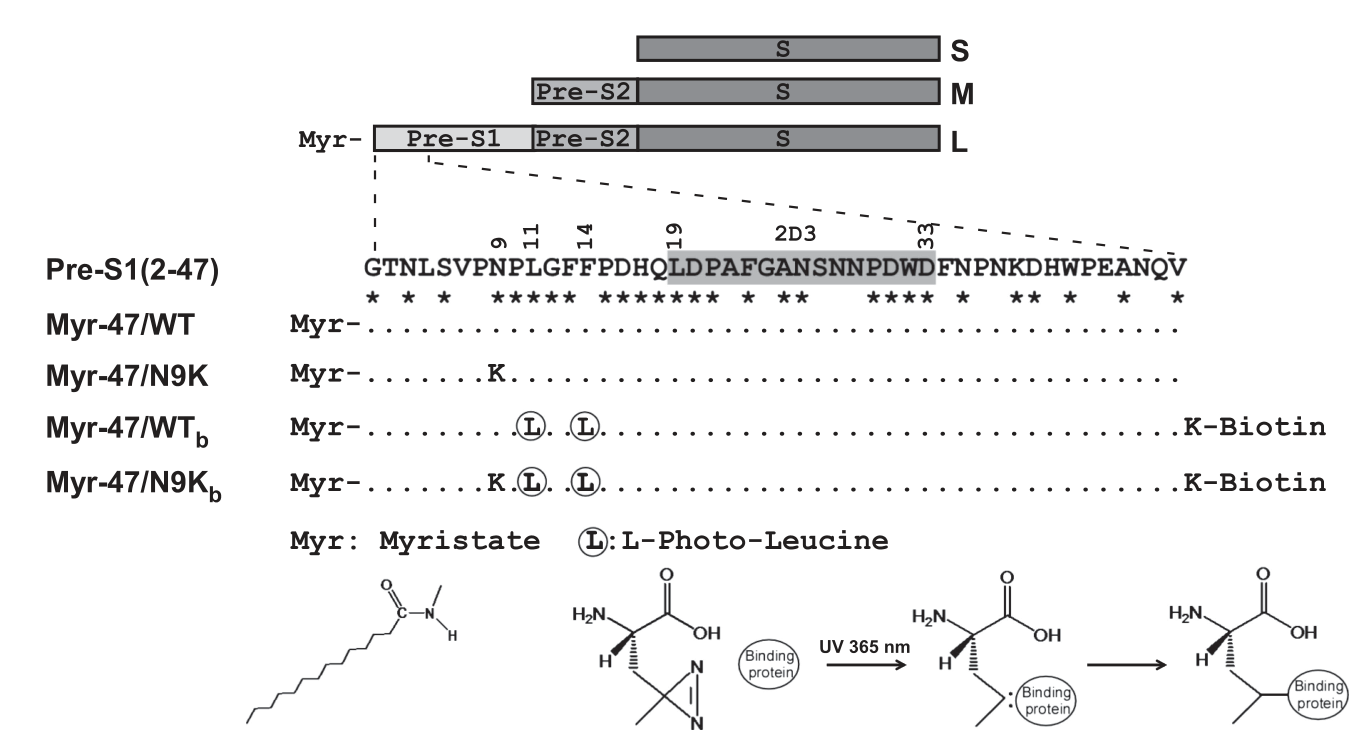
用于识别 L 包膜蛋白pre-S1相互作用蛋白的光反应配体肽
光反应配体肽在紫外光照射下可与结合蛋白发生共价连接,有助于增强pull down实验的特异性,发现与pre-S1特异性结合的目标蛋白。作为整个研究起始的基础,第一步就是要构建有效的pre-S1相关光反应肽及相应抗体。
既往研究表明,来源于L蛋白N端(pre-S1结构域的第2-48氨基酸残基)的肉豆蔻酰化多肽已经被证明可有效阻断HBV/HDV的感染过程,这提示两者可能存在竞争性抑制(即肉豆蔻酰化的多肽片段很可能与HBV/HDV在肝细胞表面有相同的受体)。
下面的结构图系我个人解读,可能有不对的地方。谨慎观看。
基于上述事实,研究者基于N端肉豆蔻酰化的pre-S1结构域的第2-47氨基酸残基(Myr-47)进行异构体的设计与合成。如下图所示,设计主要包含2个方面:
- 设计突变型多肽。具体的做法是将第9位的天冬酰胺(N)替换为赖氨酸(K)残基。一般来说,N属于极性中性氨基酸,而K属于碱性氨基酸(多一个NH3+基团),这种替换很可能会造成多肽的功能变化(甚至是Loss-of-function)。当然,生物学不是数学,需要实验进一步证实。也许既往研究已经发现这个位点的置换是有效的;也许是本研究的作者们自己试出来的。
- 设计带有
L-光亮氨酸(C5H9N3O2)基团的多肽(在第11位和第14位氨基酸残基上)。一般来说,L-光亮氨酸基团具有光反应性,在365nm波长(紫外波段)的照射下可以与结合蛋白形成稳定的共价键(是与二硫键吗?),并可被相应的实验方法所分离,因而可用于蛋白质-蛋白质相互作用(PPI)的相关研究。根据wiki的记录,Photo-leucine相应的生化技术大概是2005年前后被发明出来的。而本研究成文于2012年,估计实验是2007年以后做的,距离新技术发明只有2年左右;当时能成熟运用此技术的来研究HBV受体的团队应该并不多。我推测技术门槛也是本研究的难度之一。此外,含L-光亮氨酸多肽的C端的第48位赖氨酸(K)还连接了生物素(Biotin),我推测是为了方便通过生物素-生物素抗体的强连接,然后用IP对modified-Myr-47-目标受体复合物实施pull down,从而达到分离纯化的目的。
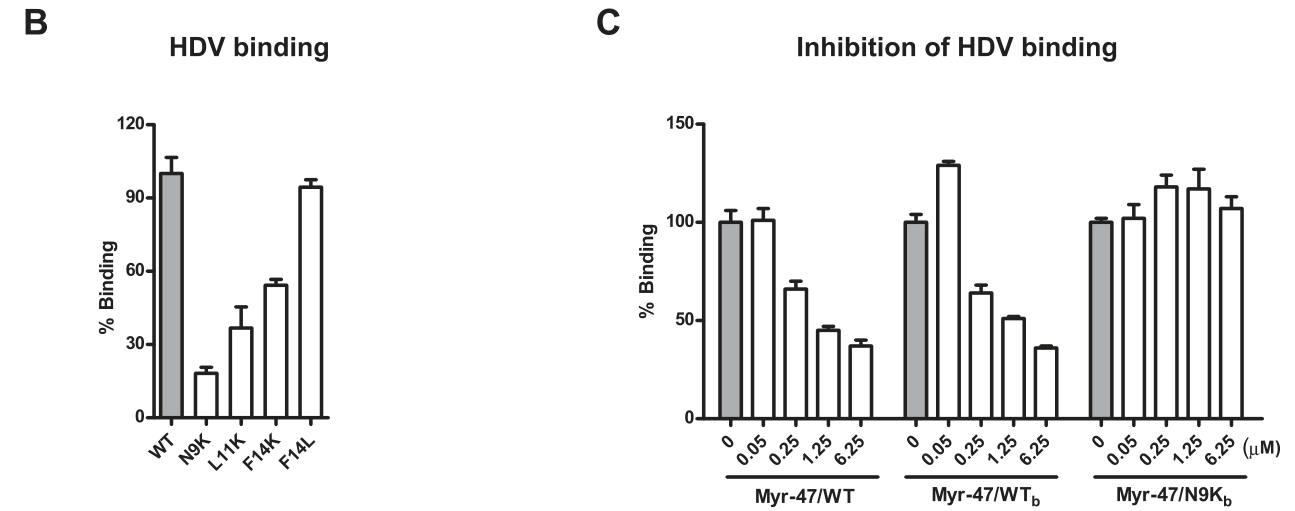
将实验多肽设计并合成后,研究者即开启了系列分子生物学实验研究它们的功能差异了。实验表明,不同HBV包膜蛋白异构体影响HDV-PTH(primary Tupaia hepatocytes, 原代树鼩肝细胞)结合的能力有所差异。其中,Myr-47/WTb 诱饵肽影响HDV-PTHs结合的效应具有剂量依赖性,而Myr-47/N9Kb(与WTb的9位氨基酸残基不同,由K代替N,见上图)对HDV-PTHs结合无显著影响,这充分表明了Myr-47/WTb 竞争性拮抗HDV的能力依赖Pre-S1结构域:
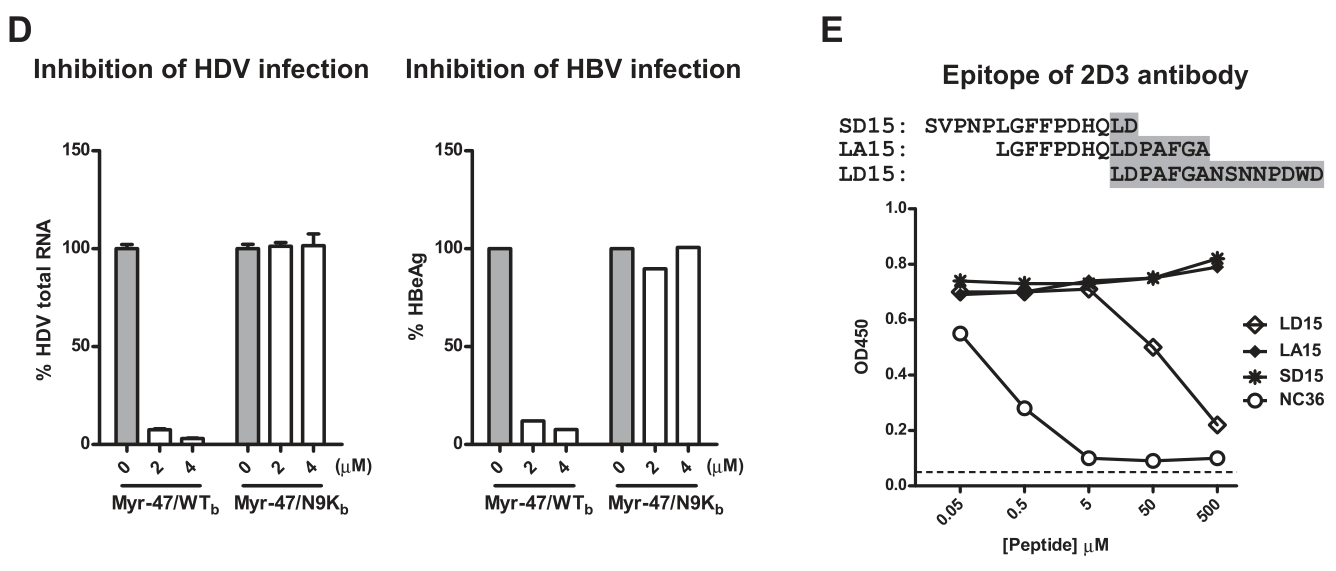
通过HDV RNA水平/HBeAg滴度的检测,进一步证实Myr-47/WTb拮抗HDV和HBV感染的作用具有剂量依赖性。为了制作pre-S1的特异性抗体,通过设计3个pre-S1段不同区域的15氨基抗原肽,ELISA实验进一步表明了anti-Myr-47/WTb抗体——2D3抗体的抗原表位在pre-S1的第19–33位残基:
鉴定NTCP为 pre-S1 的特异性结合蛋白
前面确定了光反应多肽Myr-47/WTb影响HBV/HDV-PTH的结合。下面研究者需要在原代肝细胞里确定Myr-47/WTb与哪个肝细胞受体相结合,并且鉴定出这个未知蛋白。
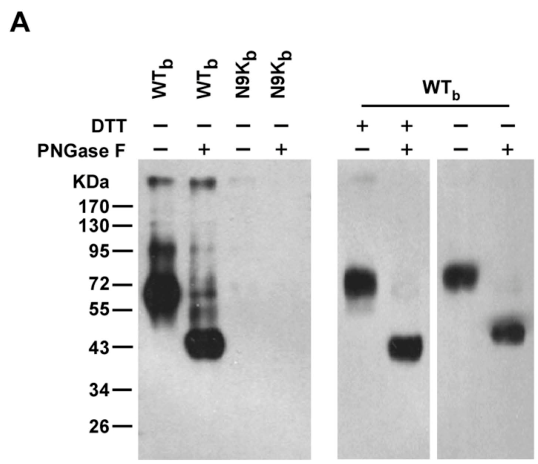
据了解,二硫苏糖醇(DTT)使免疫球蛋白发生变性、还原反应,使其轻链和重链拆分开,但不影响其原有的糖基化修饰或末端修饰;PNGase F可介导多肽的酶促糖链水解,不完全去糖基化。
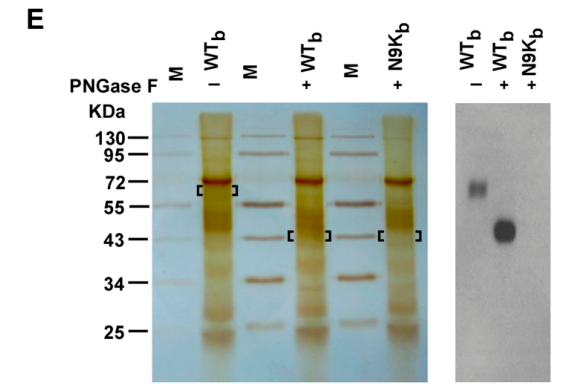
通过免疫共沉淀和Western Blot(mAb 2D3)实验,研究者成功检测到WTb-目标蛋白复合物的条带大约处于65KDa,并且经PNGase F不完全水解后与WTb结合的主要成分约为43KDa;DTT则并不能使目标蛋白水解。这表明WTb-目标蛋白复合物是高度N端糖基化的(这意味着WTb-目标蛋白复合物具有一定的PNGase F酶水解特征,这种特征有助于在后续过表达实验中进一步确认目标蛋白)。
结合竞争实验表明,Myr-47/WTb-目标蛋白复合物的相互作用会受到Myr-47/WT的干扰,却不会受到Myr-47/N9K的干扰,说明Myr-47/WTb、Myr-47/WT两者所结合的目标蛋白是一样的。
假设IP实验所体现的“条带”是Myr-47/WTb和其它蛋白(非目标蛋白)的非特异性结合。结合竞争实验可以用来确认背景噪音是否真实存在。如果噪音存在,下图WT的200、40会看到明显显影,并且8与0的对比会更不显著。而实验结果和假设推导的结果相反,说明假设不成立,即Myr-47/WTb和该蛋白的结合是特异性的。这种思想在数学上叫“反证法”。这个实验的设计还是比较巧妙的(也有可能只是一种常规操作,是我没什么见识,哈哈!)。
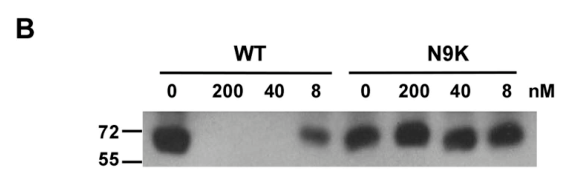
随着Myr-47/WTb在PTH培养的时间延长,Western Blot(mAb 2D3)表明PTH中的Myr-47/WTb的含量快速下降,这表明Myr-47/WTb与PTH结合具有时间依赖性(逐渐降解?):
这个实验的意义是什么?
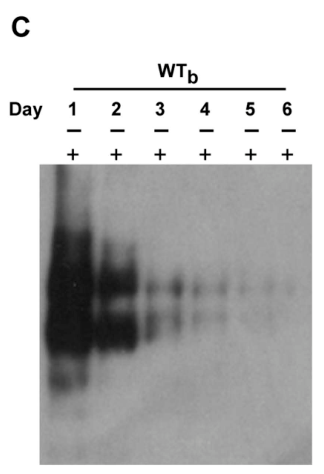
在原代人肝细胞(PHH)中,可以观察到Myr-47/WTb与目标受体结合的条带的分子量类似于PTH中的结果,这提示两者很可能共享同一/结构相似的受体:
在不同物种的细胞系进行的实验结果类似,这个证据是比较强的。实验做到这一步,可以确定大方向是对的!
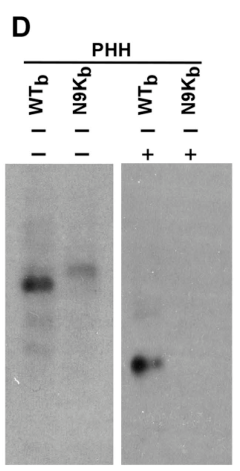
基于前面坚实的研究结果,作者选取了原始的65KDa和水解后产生的43KDa条带附近的蛋白进行MS检测:
通过与此前建立的Tupaia hepatocyte protein database的比对,发现在65KDa和43KDa处都发现了一个NTCP同工体蛋白(tsNTCP),它和人NTCP蛋白有83.9%的序列是一样的:
这个数据库是研究者们根据树鼩的转录组学深度测序确定的。这个基础能力和工作量在2012年应该算是很强的。不过在文中,作者只是轻描淡写地提了一句,附了几个图。恐怖如斯!
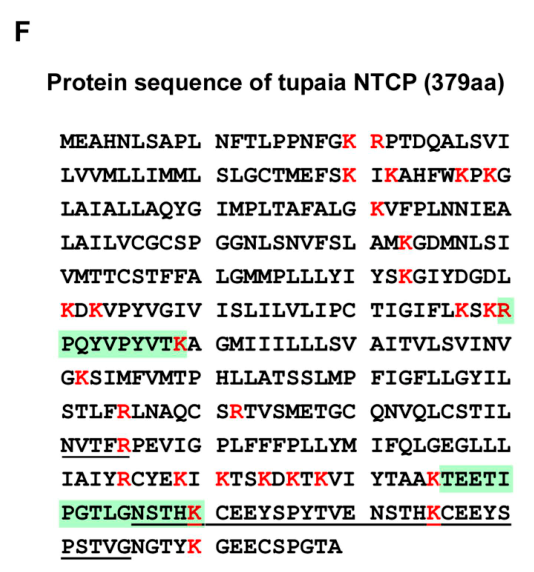
到了这一步,虽然还没有进一步论证,但NTCP是目标受体的可能性还是很大的。除了坚实的鉴定研究外,因为一般认为NTCP是参与胆汁酸循环的蛋白,与肝脏(胆汁是只有肝脏才会分泌的消化液)的功能联系十分密切;而HBV/HDV也是肝特异性病毒。这种生物学上的相似性会进一步增加NTCP是目标受体的概率。我估计,研究者们筛出NTCP的时候就知道自己的大方向是对的。这也算是分子生物学中常用的技巧之一吧,和医生诊断、警察断案的原理类似,都是基于贝叶斯公式,即通过控制先验概率增加后验概率,逐渐找到病因(疑犯)。
确认NTCP是 pre-S1 的特异性结合蛋白
前面IP取材、MS检测、序列比对等工作初步筛选了NTCP作为HBV/HDV肝细胞受体的有力侯选者,但仍需要在功能学、机制学上进一步确认(因为这时NTCP完全有可能是一个伴随的无关蛋白;特别是在生物学实验精度有限的情况下)。本节的主要目标是验证NTCP与pre-S1的结合是否是特异性的。
通过NTCP-C9(human/ts; C9属于tag)在293T细胞中构建过表达体系,研究者进一步确认,无论是human-NTCP还是ts-NTCP,NTCP-WTb相互作用是存在的,并且不耐受PNGaseF水解,分子量的变化与此前的初筛特征是一致的:
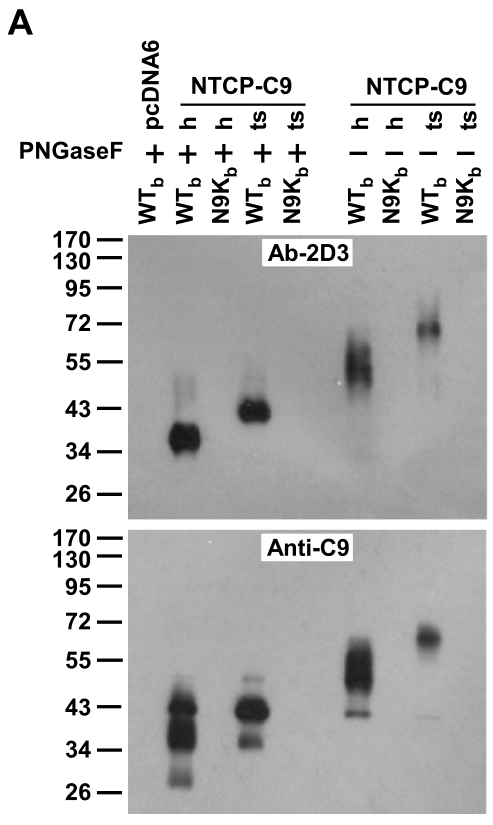
细胞免疫荧光表明,WTb可与NTCP结合并定位于肝细胞膜(左上);Myr-47/WT可竞争性结合NTCP表位,从而只有少量WTb可与肝细胞膜表达的NTCP结合(左下);N9Kb-NTCP和hSDC2-WTb均不能结合(右上/右下)。这表明Myr-47/WTb-NTCP的结合是特异性且依赖pre-S1结构域:
hSDC2-EGFP (encoding human heparan sulfate proteoglycan core protein fused with EGFP at C-terminus)
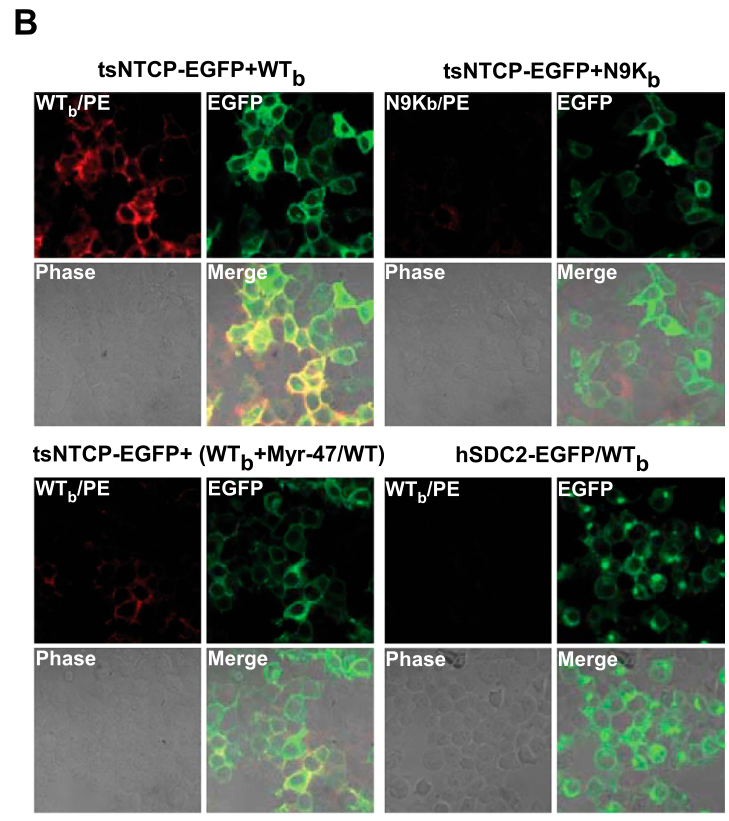
人肝癌细胞系Huh-7在自然状态下对HBV/HDV不敏感,hNTCP表达也处于较低水平。Huh-7过表达hNTCP实验 + 流式分析表明,pre-S1多肽可与hNTCP结合:
过表达hNTCP-Huh-7细胞模型估计成为HBV/HDV研究的经典模型了。毕竟肿瘤细胞可以无限增殖,研究起来可比原代肝细胞好用多了。
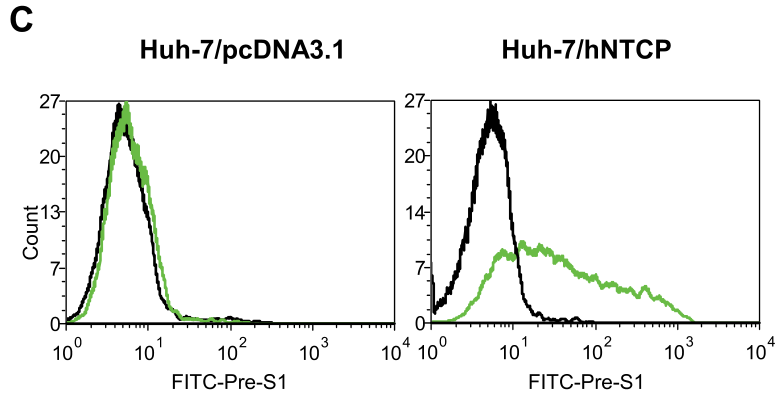
Huh-7细胞转染hNTCP或对照质粒24h后,对其进行HDV(野生型HDV或者pre-S1结构域携带N9K突变的HDV)感染。通过qRT-PCR检测结合病毒体。实验表明,tsNTCP/hNTCP转染后Huh-7细胞的结合病毒体的含量上调,这表明——NTCP可增强Huh-7感染HDV的能力,并且这个过程依赖HDV L蛋白的pre-S1结构域:
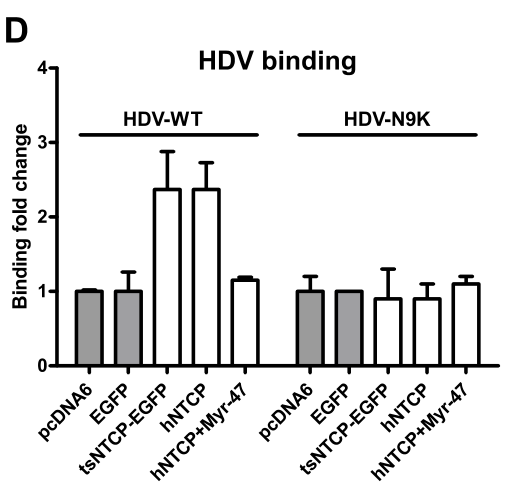
HBV 和 HDV 感染需要 NTCP 表达
前面的工作主要关注NCTP-病毒结合的关键机制。下面,研究者们需要进一步探究HBV/HDV感染的过程是否依赖NTCP,以确定其是否存在重要的生物学价值(即所谓的biological importance; 毕竟很多蛋白可能是passengers而不是drivers)。
结合tsNTCP RNA干扰实验和病毒RNA(全RNA、基因组RNA、反基因组RNA)的检测,研究者发现,与对照组相比,si-NTCP可降低PTH细胞中的HDV滴度。类似的,随着感染时间的增加(分别在感染后第6、9、12天观测),si-NTCP细胞的HBV滴度上升的幅度显著低于对照组。
AAV8-HBV是重组腺病毒相关病毒,其进入肝细胞的过程依赖AAV8衣壳而不是HBV包膜蛋白;实验表明AAV8-HBV感染对si-NTCP没有反应,这间接表明NTCP介导HBV进入细胞的过程特异性依赖HBV包膜蛋白。
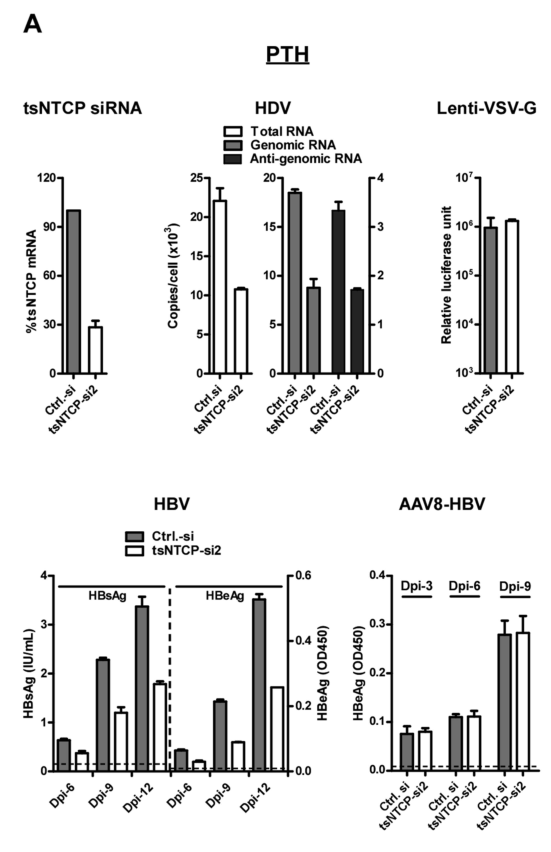
类似的,在诱导hNTCP过表达的人HepaRG肝细胞中,研究者发现si-hNTCP可抑制HDV RNA转录和逆转录。通过ELISA实验、RNA检测,si-hNTCP抑制HBV增殖的作用具有明显的时间依赖性:
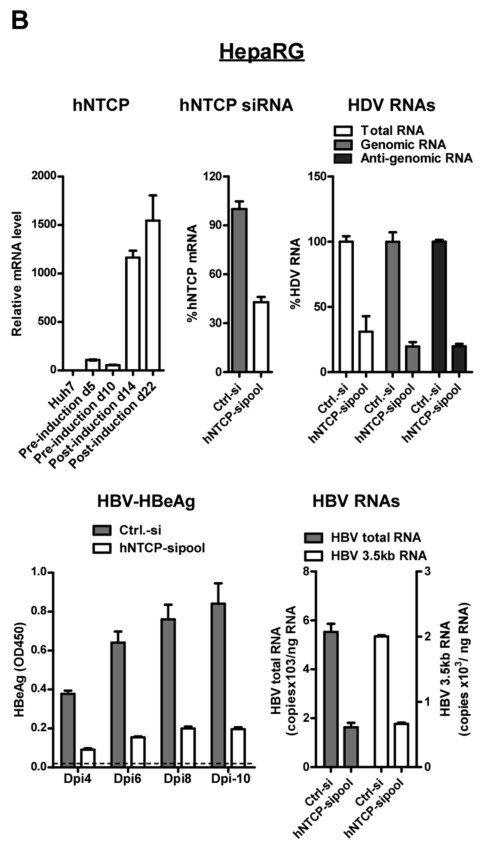
类似的,在人原代肝细胞中,与对照组相比,si-hNTCP可显著抑制HBV增殖:
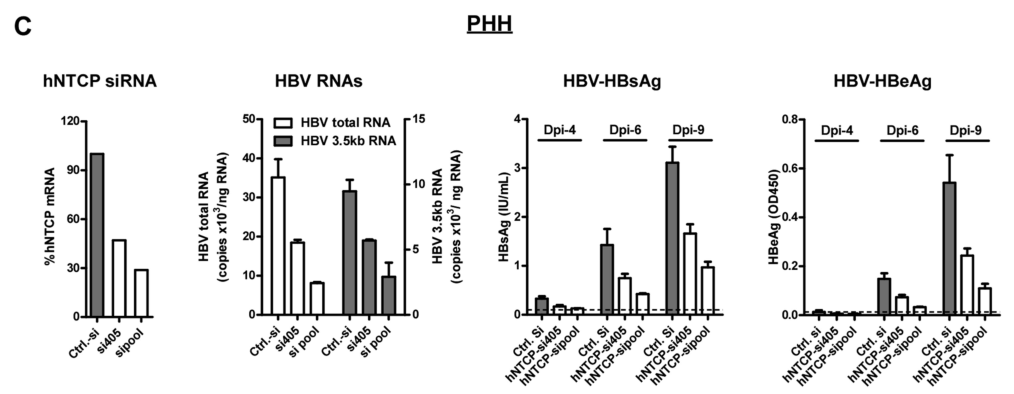
NTCP表达使不敏感的肝癌细胞易受HDV和HBV感染
前面的干扰实验已经初步证实NTCP介导HBV/HDV感染的功能。本节主要是通过功能获得实验进行更加严谨的证明。
肝细胞细胞系不同程度表达hNTCP,以PTH和PHH的水平较高,它们在自然状态下也是HDV/HBV易感细胞:
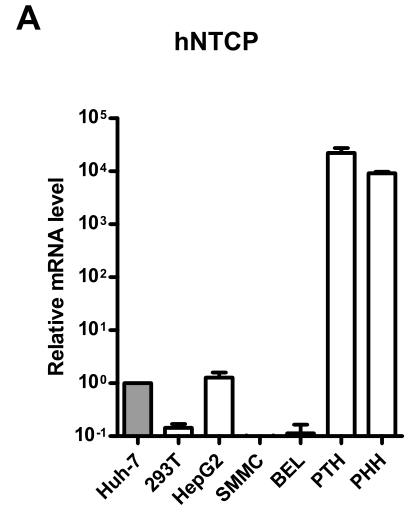
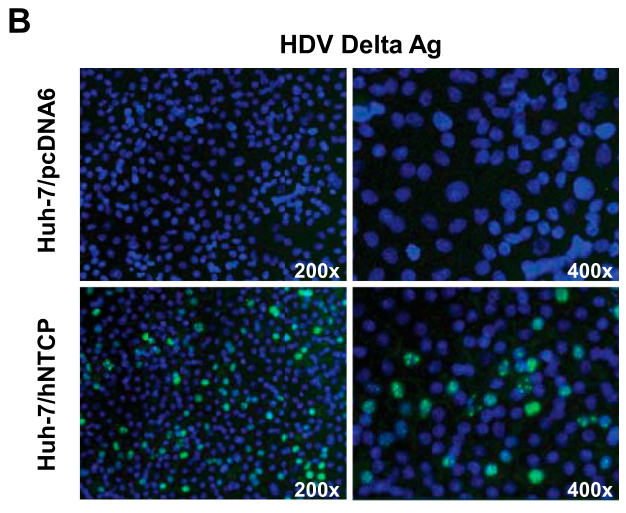
研究者发现,过表达hNTCP后,部分huh-7细胞对HDV敏感,可检测出HDV Delta Ag,这也表明NTCP是HDV感染和繁殖的关键功能分子:
左下、右下图中并不是所有的细胞都有明显的荧光,可能是转染效率有限。
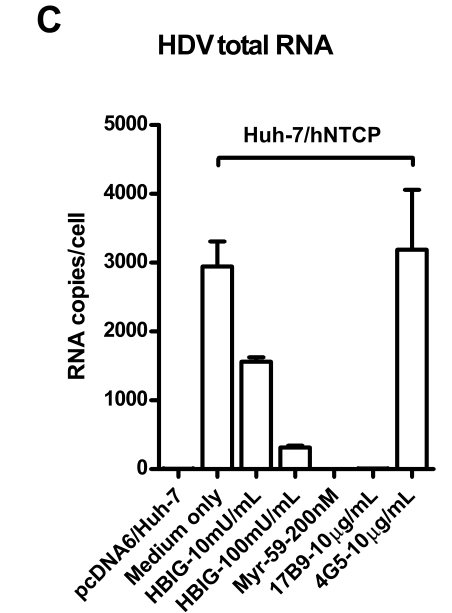
过表达hNTCP的huh-7细胞感染HDV后,HBV进入抑制剂(HBIG/Myr-59/anti-HBsAg等)的使用可抑制HDV总RNA的表达。这表明hNTCP介导HDV感染的主要过程是进入步骤:
(C) Huh-7 cells transfected with hNTCP were infected with HDV similarly as in panel B in the presence or absence of HBV entry inhibitors: HBIG (hepatitis B immune globulin), Myr-59, and anti-HBsAg mAb, 17B9. 4G5 was used as an antibody control. HDV RNA copies of infected cells were quantified by real-time RT-PCR on 6 dpi. 这是因为HBV/HDV共享类似的包膜蛋白,所以使用的是HBV进入抑制剂。
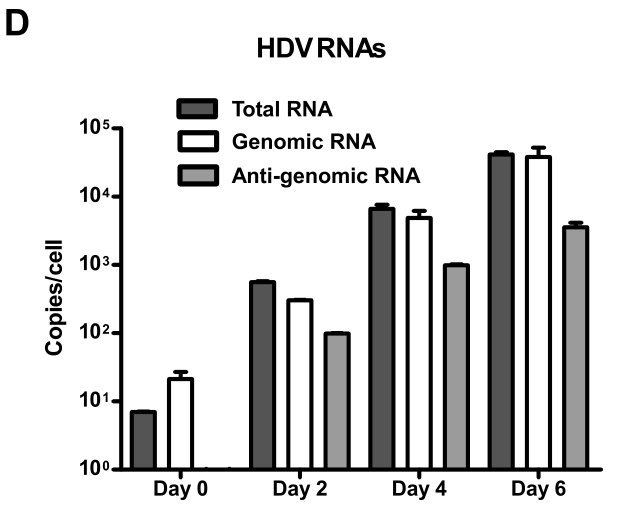
hNTCP介导HDV感染的过程具有时间依赖性:
这里的时间依赖性能体现hNTCP的功能吗?如何排除感染HDV后的自然增殖过程呢?——由于HBV/HDV是在肝细胞内进行繁殖,等宿主细胞裂解死亡后,释放出来的新病毒颗粒再次感染新的肝细胞,在这个过程中virus entry是关键步骤。从这个角度上看,病毒entry与病毒繁殖的程度应该是成正比的,观察RNA copies对于判断entry的效率是有用的。
hNTCP介导的HDV感染的过程增强还具有剂量依赖性。比如,随着HDV浓度的升高,hNTCP介导的HDV感染的过程增强:
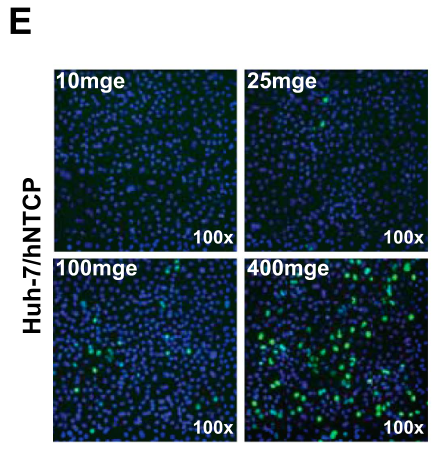
此外,随着hNTCP转染浓度的升高,hNTCP介导的HDV感染的过程增强:
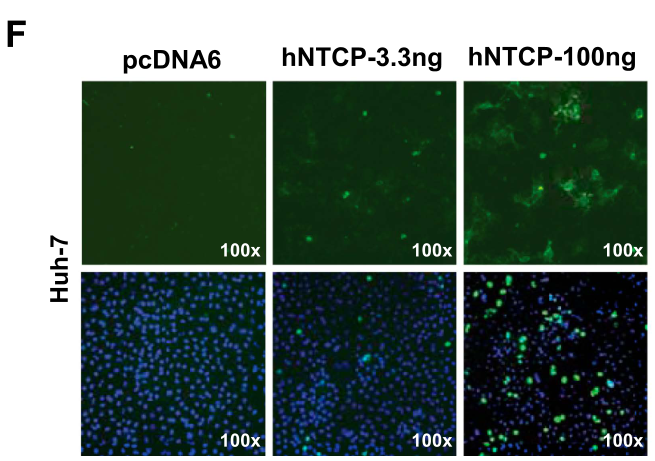
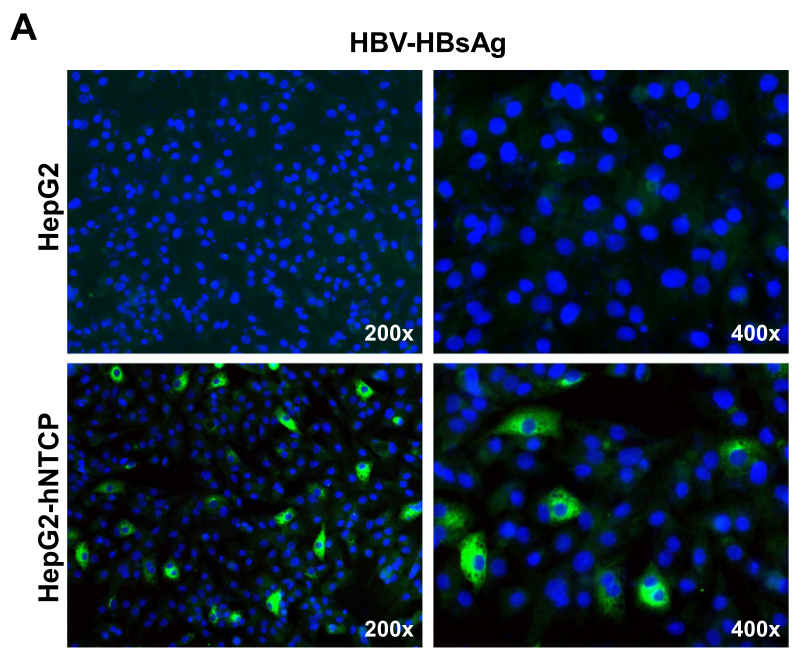
除了HDV外,作者也系统地探究了hNTCP介导HBV感染的作用。类似的,研究者们发现,在过表达hNTCP的HepG2细胞中可观察到HBV-HBsAg的表达,这表明hNTCP介导了HBV感染:
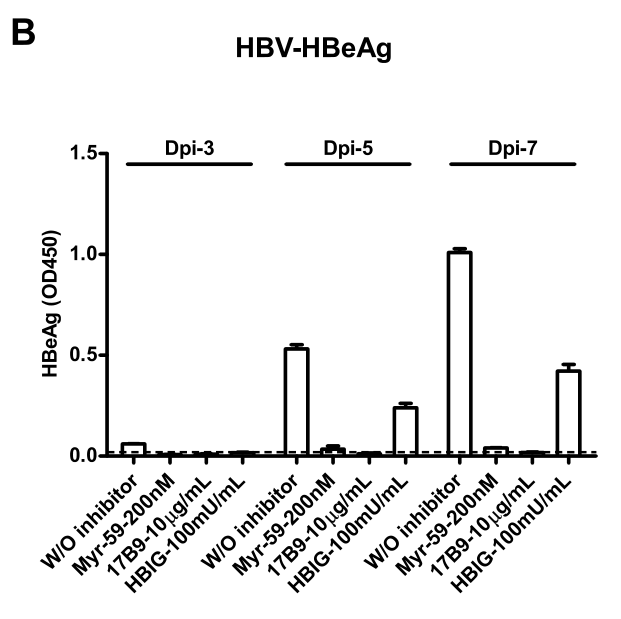
与HDV结果类似,通过检测HepG2分泌的HBV-HBeAg蛋白,研究者证明进入抑制剂可阻断HBV感染过表达hNTCP的HepG2细胞,这表明hNTCP介导HBV感染的过程发生在virus entry这一步骤:
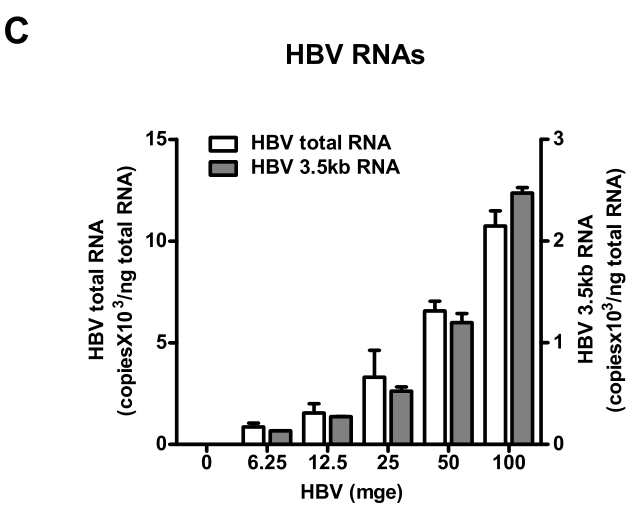
类似HDV的结果,研究者发现:随着HBV浓度升高,过表达hNTCP的HepG2细胞的HBV RNAs(包括前基因组3.5 kb RNA)水平也随之升高,体现出HBV剂量依赖性:
为什么还要检测HBV 3.5kb RNA?
(C) HBV infection efficiency is correlated with the viral inoculum dose. With increasing dose of HBV, 2 × 10 5 HepG2-hNTCP cells were infected as indicated. HBV RNAs in infected cells was examined on 10 dpi with real-time RT-PCR for the total HBV RNAs and the 3.5 kb transcripts.
HBV is a small enveloped hepadnavirus that replicates via reverse transcription of the pregenomic 3.5 kb RNA within its capsid to generate the viral 3.2 kbp partially double-stranded DNA genome [4,5]
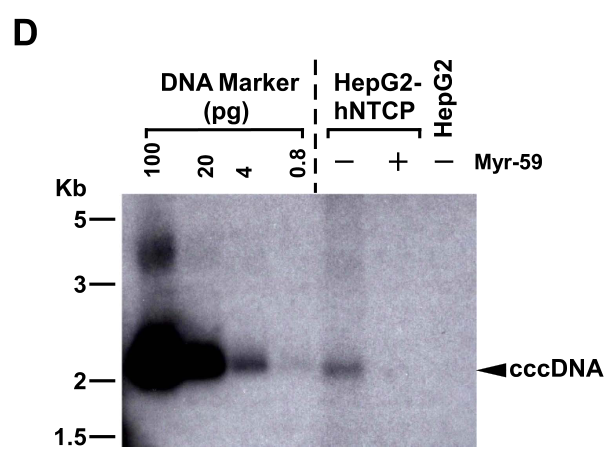
通过Southern Blot检测cccDNA,研究者发现HBV进入抑制剂Myr-59可抑制HBV在过表达hNTCP-HepG2细胞的感染性cccDNA复制(至少在0.8pg以下):
closed circular DNA (cccDNA) 闭环DNA,具有感染性
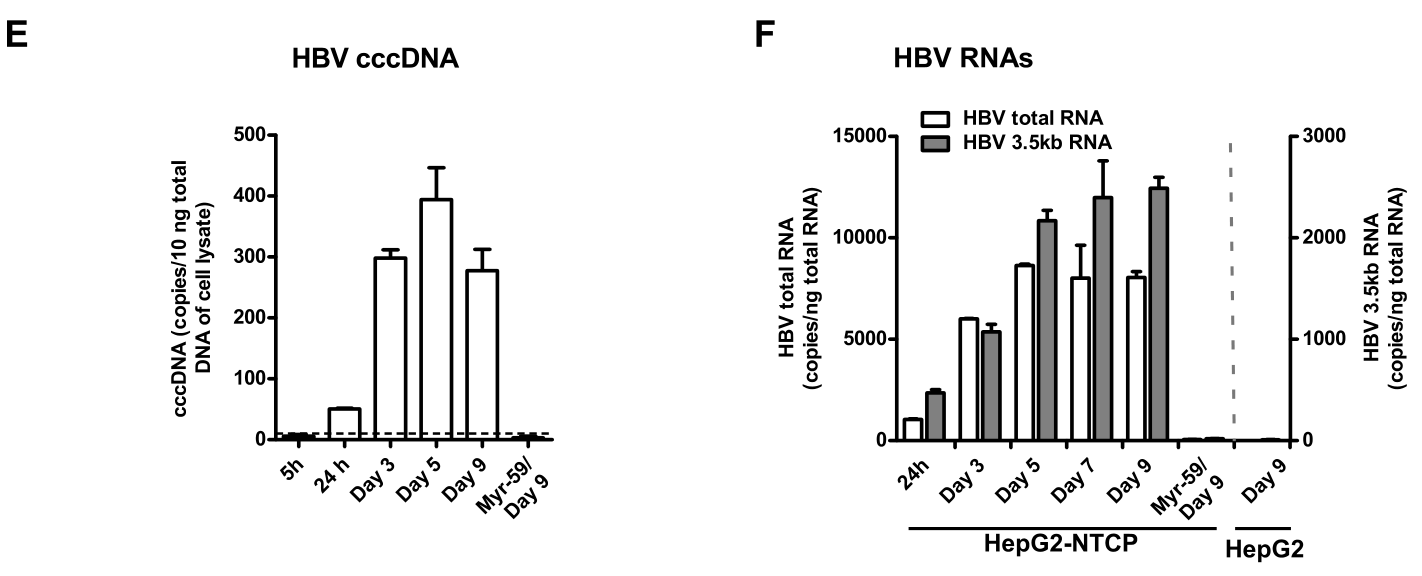
更进一步,研究者们发现Myr-59可中断HBV cccDNA的复制并促进其降解(E);Myr-59还可显著减少HBV总RNA/HBV 3.5kb至较低水平。这充分证明靶向进入过程可显著抑制HBV对过表达NTCP的HepG2细胞,也充分表明NTCP通过控制进入过程介导HBV感染HepG2,即NTCP是HBV的天然受体:
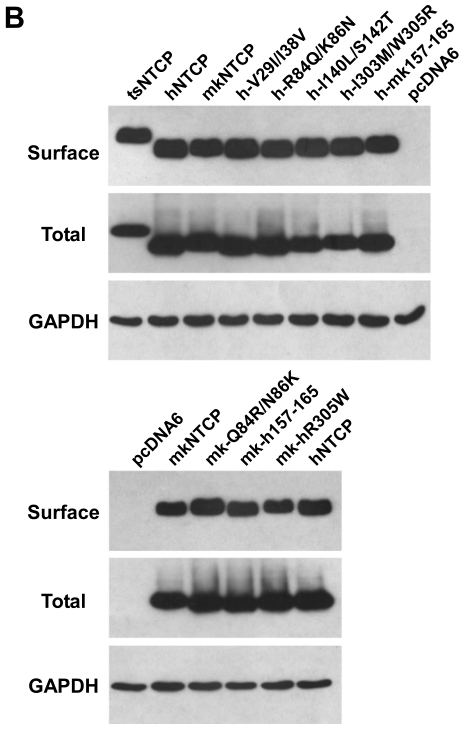
人NTCP的 157-165 残基对 pre-S1 结合和病毒感染至关重要
前面的研究已经基本明确NTCP作为HBV/HDV的功能性受体发挥作用。出于严谨,最后研究者们要找到影响NTCP功能的关键结构域(所谓知其然,也要知其所以然),为证据链增添最坚实的基础。这是分子生物学研究的基本思想:既然某个分子具有一定作用,那么一定存在决定它作用的结构。这种思想类似于物理学中证实理论的预言一样。
研究们设计了在不同物种的NTCP中的不同序列进行突变或置换以探究不同NTCP片段对其功能的影响。通过在293T细胞中过表达NTCP的同源蛋白或者异构体,发现hNTCP的第157-165片段是影响其与pre-S1结合的关键结构:
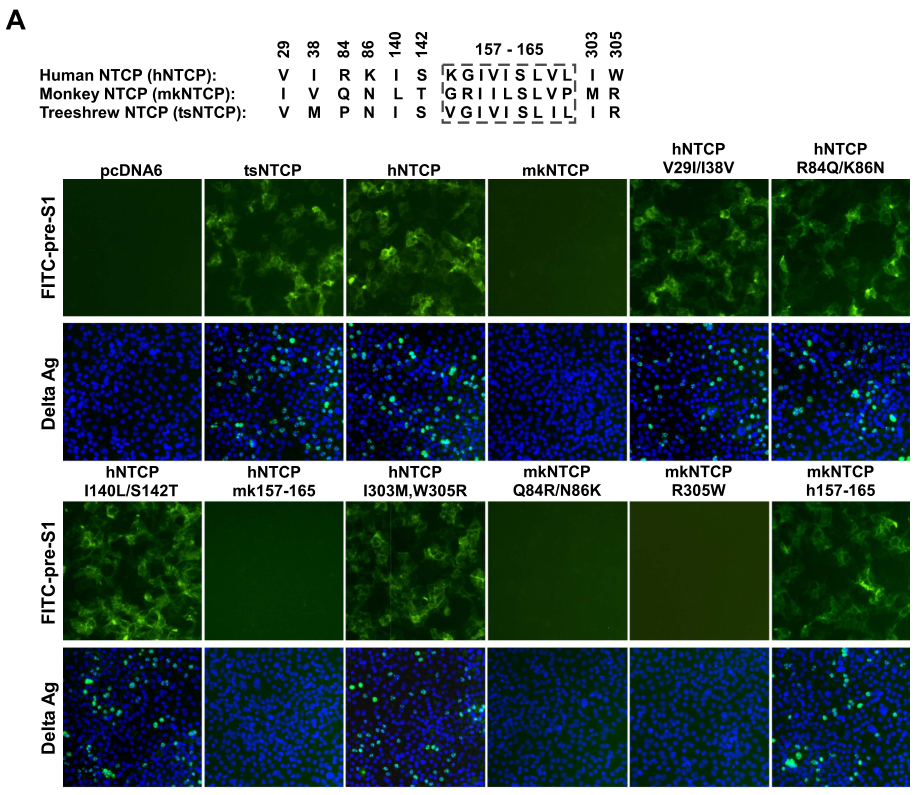
类似的,通过HBVeAg和HBV 3.5 kb RNA的测量,发现hNTCP的第157-165片段是影响HBV进入肝细胞并繁殖的关键结构:
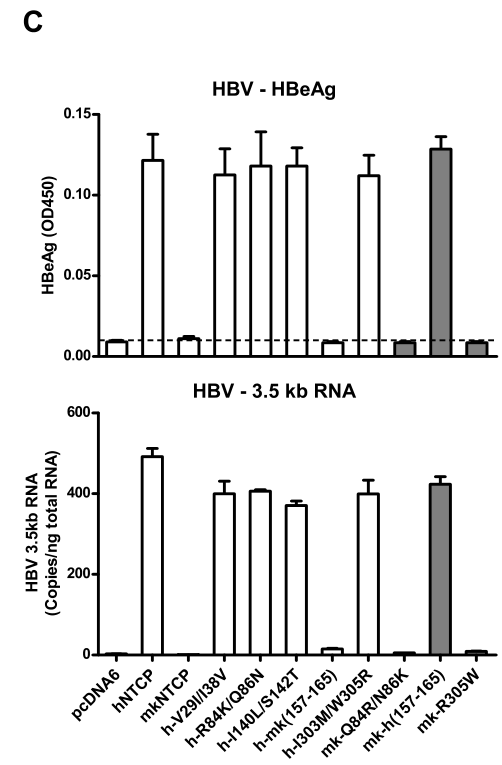
通过全蛋白/膜蛋白的Western blot实验,发现各种NTCP同源蛋白/异构体均主要表达在细胞膜上:
小结
本文的研究思路清晰,论证过程如行云流水,一气呵成,也没有特别难懂的地方,阅读的过程还是十分享受的。控制变量的技巧包括:敲低/过表达 vs. Control、多细胞系/跨物种细胞系、结合竞争实验、失活突变 vs. WT、时间/剂量依赖性等,均是基本的分子生物学实验的设计方法。附图我没细看,但只看正文基本上可以了解本文的研究框架和研究内容了。基于“本研究是一项优秀的分子生物学研究”这一事实,我谈几点自己的体会,与大伙共勉:
一、研究有攻克希望的重大问题。有一些问题很重大,但是短期之内似乎没有解决的希望;如果选择研究这种问题,很可能一直没有产出,那么在科研的道路上就很难走下去;如果选择一些不重要的问题,就算很快做出来,学术影响力也很极其有限,那么在科研的道路上就很难走得更远。“选择比努力更重要”。这是一个非常重要的点,也是很考量研究者大智慧、大能力的点。恰到好处的选题本身也是一项高水平的工作。如果去看大科学家们的传记,这种例子在历史上比比皆是。本研究的成功有几个重要前提:Pre-S1的作用已经较确切;相关技术已经被开发出来,正处于推广阶段;模式动物选择有限,研究门槛相对较高;模式动物转录组数据在手,序列比对工作基础已打牢。其实,之前不少团队都探索过HBV/HDV的天然受体,但均以失败告终,说明选题难度很大,一定程度上也阻拦了不少后来者的持续跟进,竞争者少一些(但都是有力竞争者)。从前面分析的几点来看,就算李的团队不做,我估计再过个几年也会有人做出来的,这是技术和流程成熟的必然结果。不过,结合敏锐的科研嗅觉、扎实的技术基础、不怕困难的精神态度,再加上稳定的基金支持,最后是李最早率领团队完成最关键的一步,确实不容易,也是值得称赞的。这种选题的经验和艺术很值得年轻学者学习。由于中国科研绩效考核制度的实施,许多年轻学者都不得不选择短平快的课题做,长此以往固然对国家发展不利,自身发展也会受到很大限制的。
二、逻辑高于一切。在选题完成后,本文的论证工作是以逻辑为导向的,层层剥茧,从而实现完整、严谨的论证。以现在的眼光看,文章的图谈不上美观;数据量也谈不上巨大。但这仍是一项十分优秀的研究。做科研,逻辑通顺、证据充分是最重要的。如果图片美观/海量数据是重要的证据,那么它们就是必需的;否则只是锦上添花(甚至是起迷惑审稿人的作用)的东西罢了。但平时接触到不少人的观点,将美观/数量当作是好研究的关键特点,我觉得是很不妥的。研究者应该将精力花在选题上;确定问题重要且可行性强的前提下,根据目标制定实验方案,需要多少数据准备多少数据,没必要堆砌数据。论文的图表可以说明对应的问题即可,美观不美观其实无关痛痒。再说,审美其实是一种非常主观的东西,以外观定好坏其实是很武断的。就像评论一个人看TA的外貌一样,虽然没有大错,但是终究是比较肤浅的。以问题为导向,而不是外观/数据量为导向,才是一种端正的科研态度。
三、做别人可以follow的研究。本研究完成之后,提供了很多平台性的东西供后人所用,比如过表达NTCP的Huh-7细胞作为HBV/HDV体外感染模型。这一点其实和第一点有重合之处。毕竟你解决了一个重要问题,后面的人自然很容易在你的基础上深化该领域了!
除了这些和科研有关的心得外,我也从本文感受到了大自然的神奇之处。显然,人类与HBV的共同演化可能存在很长时间了,甚至是在人与类人猿的祖先还没进一步分开的时候。树鼩与人类的关系相对较远些,这可能是一种趋同演化。从策略上看,HBV其实是一种很“成功”的病毒,发生了恰到好处的变异,和人类肝细胞的NTCP苟在了一起;并且通常不在生育年龄前造成严重的急性疾病,这样宿主就有机会将他的基因传给后代,从而避开自然选择的压力。有时看这种特异性疾病谱,内心还是很震撼的。这种特异性完全是由内在分子结构所决定的,完全是客观规律,不随人的意志而改变。揭开这种大自然秘密的人,其内心之爽快,也是很难令人想像的。所谓的“科研的快乐”,差不多就是这样吧!有时候看科学史,发现很多卓越的科学家们的生活都是很艰难的,但他们常常还是忍受着常人无法想像的痛苦完成他们的研究。我有时在想,也许他们不是崇高,只是对发现这种东西上瘾罢了。同时也庆幸自己生在一个重视科学的年代,在某些条件下,科学家可以心无旁骛地做研究而不用关心其它问题。当然,现实是,营造良好的科研环境,我们还有很多路要走呢!
总之,这是一篇非常优秀的论文,常看常新。也许,这就是经典之作的魅力吧!
扩展阅读
- A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus–induced lung injury | Nature Medicine:2003年SARS病毒的人类受体是ACE2(2019年的SARS-Cov-2也是它)。
---------------
完结,撒花!如果您点一下广告,可以养活苯苯😍😍😍
